Cell Membranes Fine-Tuned for Transport

It goes without saying: immigration is one of the most contentious political issues today. In view of the animus surrounding the undocumented people within the US borders, some might find it surprising to learn that the immigration process into the US is based on a set of principles with which few would disagree.
These principles are:
- Reunite families
- Admit people with value to the US economy
- Promote diversity
- Protect refugees
Ultimately, the US immigration policy is centered around fairness. The Immigration and Nationality Act (INA) allows 675,000 permanent visas to be issued each year across all visa categories. Consistent with the INA’s principles, no limit is set on the number of permanent visas that can be granted for spouses, parents, and children under 21 of US citizens.
No one country can be granted more than 7 percent of the total visas issued. Also, caps are placed on the different types of permanent and temporary visas that are issued.
In other words, the US has a well-thought-out immigration strategy, with the number of visas issued each year carefully adjusted in light of a set of well-founded principles.
Border Control in Cells
Like immigration officials, cells pay close attention to their borders. Cells rely on their membranes to regulate the numbers and types of molecules that move into and out of the cell or into and out of organelles within the cell. This movement is usually mediated by proteins embedded in cell membranes. Much of the movement of materials across cell membranes is highly selective, with each type of transporter protein ferrying specific molecules while excluding others. Some proteins actively transport specific materials into and out of the cell against the concentration gradient (from low concentration to high concentration) in a process called active transport. Other materials move selectively across cell membranes through specialized protein pores. These pores don’t actively move materials across cell membranes. Instead, they allow these materials to migrate passively across cell membranes along the concentration gradient (from high concentration to low concentration). This process is called facilitated transport.
Some materials enter and exit the cell by moving through the matrix of the cell membrane in a process called passive transport. These materials tend to be low in molecular weight, uncharged, and nonpolar. They move passively across cell membranes with the concentration gradient. The cell membrane matrix forms a semipermeable boundary, however, prohibiting the movement of both high and low molecular compounds that are polar or charged.
The passive transport of materials through cell membranes carries particular importance for researchers working in biotechnology who are looking to use bacteria and yeast to produce materials with biomedical or industrial uses. If the materials produced inside the cells by genetically engineered metabolic processes have the right physical properties, researchers hope to take advantage of passive diffusion to move these compounds from inside the cell into the growth medium, where they can be readily recovered.
Unfortunately, the experimental approaches and models used to predict the flux of materials across cell membranes by passive diffusion don’t always yield reliable results. In an attempt to develop a better understanding of passive diffusion, a team of biochemists from Germany has examined the effect of the lipid components of cell membranes on passive transport.1 Their work has important scientific implications. It also can be used to marshal a case that the compositional makeup of cell membranes reflects a Creator’s intentional design.
But first a bit of background on cell membranes. Those of you who are familiar with the biochemistry of cell membranes can skip to Lipid Composition Influences Passive Diffusion.
The Biochemistry of Cell Membranes
Cell membranes are comprised of lipids and proteins. Lipids, a structurally heterogeneous group of compounds, share water insolubility as a defining property. Additionally, lipids dissolve readily in organic solvents. Cholesterol, triglycerides, saturated and unsaturated fats, oils, and lecithin are examples of familiar lipids.
Membrane Lipid Structures
Phospholipids are the cell membrane’s major lipid component. Their molecular shape roughly resembles a balloon with two ropes tied to it. Biochemists divide phospholipids into two regions that possess markedly different physical properties. The head region, corresponding to the “balloon,” is soluble in water, or hydrophilic (“water-loving”). The phospholipid tails, corresponding to the “ropes” tied to the balloon, are insoluble in water, or hydrophobic (“water-hating”).
Chemists refer to molecules such as phospholipids that possess distinctly different solubility domains within them as amphiphilic (“ambivalent in its likes”). The amphiphilicity of soap and detergents gives these compounds great economic importance.
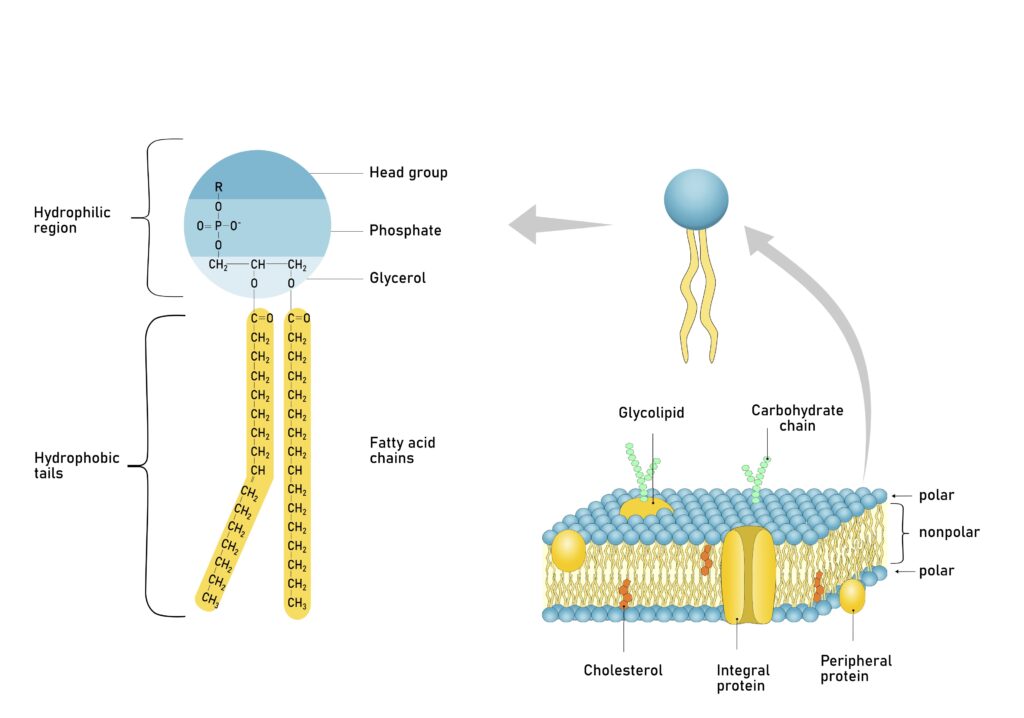
Figure 1: Phospholipid Molecule
Credit: Shutterstock
Amphiphilicity also has great biological importance. Phospholipids’ “double-minded” solubility properties play the key role in cell membrane structure. When added to water, phospholipids spontaneously organize into sheets that are two molecules thick, called bilayers. The phospholipid “heads” align adjacent to one another, and the phospholipid “tails” pack together closely. These monolayers, in turn, come together so that the phospholipid tails of one monolayer contact the phospholipid tails of another monolayer. This tail-to-tail arrangement ensures that the water-soluble head groups contact water, and the water-insoluble tails avoid water.
Phospholipids possess a wide range of chemical variability. Phospholipid head groups typically consist of a phosphate group bound to a glycerol (glycerin) backbone. The phosphate group, in turn, binds one of several possible compounds that vary in their chemical and physical properties. Phospholipids are identified by their head group substituents. For example, when choline binds to the phosphate group, biochemists refer to it as a phosphatidylcholine.
Phospholipids vary in tail length and structure. Phospholipid tails—typically long, linear hydrocarbon chains—also link to the glycerol backbone. The phospholipid hydrocarbon chains are commonly 16 to 18 carbon atoms long. Sometimes a permanent kink exists at some point along one or both hydrocarbon chains due to the presence of a double bond.
The precise mixture of the cell membrane’s phospholipids affects its physical, chemical, and, consequently, biological properties. For example, cell membranes composed of phospholipids with short hydrocarbon chains or kinked hydrocarbon chains possess a liquid-like interior. On the other hand, cell membranes formed from phospholipids having longer hydrocarbon chains without kinks have solid-like interiors.
Membrane Proteins
Proteins comprise the other major class of biomolecules that play a role in cell membrane structure and function. The cellular machinery builds proteins by joining smaller subunit molecules, amino acids, in a head-to-tail manner. Cells use 20 different amino acids with varied chemical and physical properties to form proteins. The amino acid chains that make up proteins adopt complex and precise three-dimensional structures. A protein’s three-dimensional structure determines its functional and/or structural role in the cell.
Proteins associate with the cell membrane in a variety of ways. Some, called peripheral proteins, bind to the inner or outer membrane surfaces. Others, called integral proteins, embed into the cell membrane. Some integral proteins embed only partially into the membrane interior, others penetrate nearly halfway into the membrane’s core, and still others span the entire membrane.
Membrane proteins perform different functions. Some proteins serve as receptors, binding compounds that allow the cell to communicate with its external environment. Some catalyze chemical reactions at the cell’s interior and exterior surfaces. Some proteins shuttle molecules across the cell membrane. Others form pores and channels through the membrane. Some membrane proteins impart structural integrity to the cell membrane.
Fluid Mosaic Model
Since the early 1970s, the fluid mosaic model has provided the framework to understand membrane structure and function. This model views the phospholipid bilayer as a two-dimensional fluid that serves as both a cellular barrier and a solvent for integral membrane proteins. The fluid mosaic model allows the membrane proteins and lipids to freely diffuse laterally throughout the cell membrane. Beyond the bilayer structure and asymmetry, the fluid mosaic model attributes no structural and functional organization to cell membranes.
In recent years, scientists have revised the fluid mosaic model. Instead of diffusing freely in the phospholipid bilayers, most proteins find themselves confined to domains within the membrane. Other proteins diffuse throughout the membrane, but instead of moving randomly, these proteins move in a directed fashion. Phospholipids, too, organize into domains with certain phospholipid classes laterally segregated in the bilayer. Bilayer fluidity also varies from region to region in the cell membrane.
Lipid Composition Influences Passive Diffusion
Using a combination of experimental measurements and molecular simulations, the German research team systematically studied the passive diffusion of a variety of small molecules through model cell membranes comprised of differing lipid compositions.
They learned that varying phospholipid head groups had very little effect on the extent and rates of passive diffusion. On the other hand, the phospholipid fatty acid composition did have a significant impact on passive diffusion. Specifically, the length and the degree of unsaturation (the number, type, and location of the double bonds) of the hydrocarbon chains that made up the phospholipids’ fatty acids regulated passive transport across the model membranes.
Phospholipids with longer hydrocarbon chains form thicker membranes with reduced fluidity. Both effects limit passive diffusion. Phospholipids that have fully saturated hydrocarbon chains (lacking any double bonds) also display reduced passive diffusion because the bilayers are less fluid. Phospholipids with unsaturated hydrocarbon chains form fluid bilayers that accelerate the rate of passive diffusion.
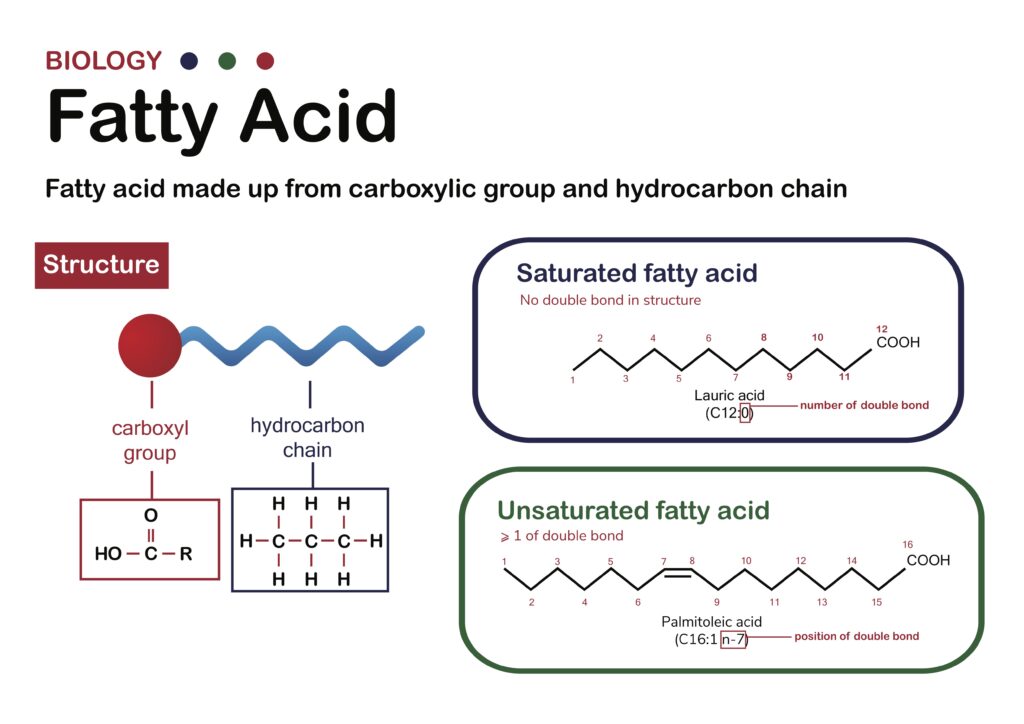
Figure 2: Fatty Acid Diagram
Credit: Shutterstock
Adding a class of lipids called sterols to the model membranes also impacted passive diffusion. As it turns out, these compounds influence bilayers’ fluidity. Depending on the identity of the specific sterol, bilayer fluidity either increases or decreases. The identity of the sterol also influences the rate of fluidity change as sterol concentration increases. Some sterols alter bilayer fluidity smoothly and continuously as their concentration in the bilayer fluctuates. Others cause more dramatic and sudden changes in bilayer fluidity as their concentration changes.
In other words, the cell can regulate the passive diffusion of small molecules by fine-tuning the lipid composition of its membranes.
The research team from Germany points out that this new insight into passive diffusion has explanatory power. It accounts for differences in small molecule permeability through the cell membranes of different microbes such as bacteria and yeast. It also explains the differences in passive diffusion observed for plasma membranes and organellar membranes within eukaryotic cells.
As mentioned, this discovery can be exploited by scientists working in biotechnology to readily harvest drugs and other compounds produced by genetically engineered bacteria or yeast. The German researchers also think it can be used to design drugs that can efficiently diffuse into target cells via passive diffusion, with the drug’s physical properties matched to the target cells’ lipid composition.
Cell Membranes and the Case for a Creator
Many biochemists hold the view that cell membranes originated through an evolutionary process that was unguided and undirected. Many life scientists regard the evolutionary process as historically contingent, predicated on a sequence of chance events producing outcomes that are ultimately happenstance. As evolutionary biologist Stephen Jay Gould pointed out in his book Wonderful Life, if we could somehow turn back the hands of time and let evolutionary history reoccur the outcome would be different, each and every time.
If this type of evolutionary process generated cell membranes, then its compositional complexity shouldn’t necessarily have any rhyme or reason to it. It’s merely what the evolutionary process cobbled together over time. Yet, the German biochemists’ research reveals that the fatty acid composition of the phospholipids that make up the bilayer and the types and concentrations of sterols in cell membranes serve at least one purpose; namely, regulating passive transport across the bilayer. In other words, a rationale undergirds the compositional makeup of membrane bilayers even though it is characterized by a bewildering complexity of lipids. This complexity does not result from a historically contingent evolutionary process. It appears to be intentional and purposeful. And, if the lipid composition of cell membranes is purposeful and based on a rationale—a molecular logic, of sorts—then, at minimum, it is reasonable to think that these biochemical systems have been designed by a Mind.
The German researchers’ study also revealed another provocative feature regarding the lipid composition of cell membranes. It appears to be exacting and precise. It is fine-tuned to regulate the flux of small molecules through the bilayers of cell membranes. Their research indicates that adjusting the fatty acid distribution of phospholipids and the sterol levels will change the rate and extent of passive transport. Clearly, cells employ this mechanism to control the flux of small molecules across cell membranes, given the difference in passive diffusion of small molecules across the plasma membranes of bacteria and yeast. And, of course, precision and fine-tuning are also hallmarks of systems that have been designed intentionally.
To put it another way, the study by the German biochemists reveals the precise features we would expect to see if cell membranes—and hence, life itself—were designed by a Creator.
Resources
- The Cell’s Design: How Chemistry Reveals the Creator’s Artistry by Fazale Rana (book)
- Fit for a Purpose: Does the Anthropic Principle Include Biochemistry? by Fazale Rana (book)
- “Biotic Borders: Cell Membranes under Scrutiny” by Fazale Rana (article)
- “Bringing Order to the Case for Intelligent Design, Part 1” by Fazale Rana (article)
- “Bringing Order to the Case for Intelligent Design, Part 2” by Fazale Rana (article)
- “Fatty Acids Are Beautiful” by Fazale Rana (article)
Check out more from Reasons to Believe @Reasons.org
Endnotes
- Jacopo Frallicciardi et al., “Membrane Thickness, Lipid Phase and Sterol Type Are Determining Factors in the Permeability of Membranes to Small Solutes,” Nature Communications (March 25, 2022), doi:10.1038/s41467-022-29272-x.